 |
||
|
|
||
| / Вне рубрик / < Вы здесь |
|
Теоретические аналитические конструкции в системе научного знания
Позитивистская интерпретация иерархий таксонов Дата публикации: 16 Мая 2001 Трудности в идеализации предметной области, как и в науке в целом, приводят к тому, что вместо теоретических миров в функции объяснения выступают их суррогаты, такие как эволюционные и редукционистские построения, замещающие и вытесняющие позитивное научное знание на периферию социальной жизни науки. Как уже отмечалось ранее, если не принимать во внимание деятельность исследователей в рамках итерации цикла таксономии, то результаты итераций можно представить в виде иерархии таксонов, существующей самой по себе, что и зафиксировано в учебных пособиях, иллюстрациях к определителям, а также во многочисленных научно-популярных работах. Онтологизация структуры определителей, попытки представить результаты описывающей и упорядочивающей деятельности исследователей как состояние, свойственное самому миру, во многом детерминируют структуру биологического (и не только биологического) мышления. Более того, логика строения биологических определителей и работы с ними в XIX-XX вв. стала рассматриваться как отображение процесса эволюции форм жизни, и отношения сходства-различия особей по признакам, формирующим цикл таксономии, выступают в эволюционных мифологемах как отношения схождения (конвергенции) и расхождения (дивергенции) признаков в эволюционном процессе. В эволюционных теориях отношения между таксонами в системе таксонов рассматриваются как отображение эволюционного процесса. Основу этих учений составляет интерпретация отношений сходства, в которых таксоны вводятся в систему, как отношения родства и происхождения. В рамках развиваемых представлений о структуре знания теоретико-эволюционные надстройки над онтологией можно рассматривать как следствие неразличения (отождествления) теоретического и эмпирического уровней исследований. Особи при этом становятся неотличимыми от таксонов и типов, а аналитические объекты отождествляются с архетипами. Естественно, что при этом отношения между особями отождествляются с отношениями между таксонами, а частным (типологическим) конструкциям аналитического знания присваивается статус всеобъясняющих теоретических миров-архетипов. Ньютоновская и эйнштейновская механики выдаются позитивистами за идеальные модели объясняющих теорий. Из табл. 3 видно, как плоский позитивистский мир весьма ограничен в своих возможностях собственной онтологией. 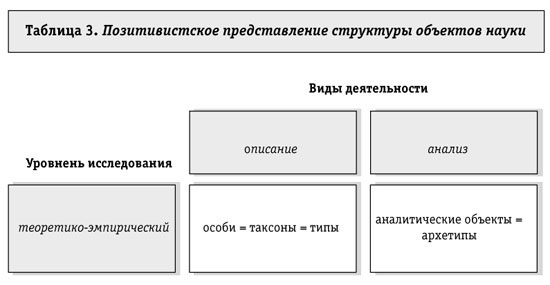 Отношения родства и происхождения, в отличие от сходства, - естественные отношения между особями и популяциями. Отношения сходства вводятся исследователями-систематиками для формирования таксонов и являются искусственными отношениями классификационной системы, адекватность которых проверяется в других отношениях таксономии. Рассматривать отношения родства как отношения между таксонами в иерархии (или другой системе) таксонов, даже если отвлечься от естественной природы родства, в принципе нельзя, т.к. таксоны - теоретические объекты, которые не могут происходить друг от друга, если только они не включены в какую-либо систему объективного идеализма, вроде гегелевской. Теоретические объекты лишены способности размножаться или происходить друг от друга без участия людей, у них нет для этого соответствующих органов. Теоретические объекты имеют потомков и предков только в очень узком смысле этого слова. В отношениях происхождения и родства могут находиться только особи. Ощущение несоответствия (здесь не приходится говорить о понимании, так как его нет) между системой таксонов и ее интерпретациями в эволюционных учениях есть практически у каждого исследователя-систематика, интересующегося филогенией. Проблема теоретических структур таксономии очень болезненна и провоцировала множество мировоззренческих и философских дискуссий. Тем не менее эволюционные концепции, основывающиеся на иерархии таксонов, должны быть исследованы на непротиворечивость, поскольку внутренняя несогласованность и противоречие известным фактам в построениях, претендующих на роль объясняющих теорий, чреваты опасными для науки последствиями. Итак, теоретические схемы таксономии во многих, если не в большинстве, случаях, основаны на установлении между таксонами отношения "происходить от" и на интерпретации отношений сходства как отношений родства: "...все существующее многообразие форм жизни возникло в результате прогрессирующего расхождения в процессе биогенеза. Хотя в целом верно, что потомком мыши будет мышь, а потомком человека человек, время от времени возникают отклонения, которые задним числом можно рассматривать как источник новых видовых форм. Эволюционная гипотеза неотъемлемо входит в основу основ способа мышления в биологии. Только эволюционная гипотеза придает смысл несомненной взаимосвязи организмов, явлениям наследственности и путям развития. Биолог может мыслить только эволюционно - другой альтернативы у него нет"89. Как известно, варианты эволюционных учений различаются, и весьма кардинально, тем, как они интерпретируют появление новых форм жизни, исчезновение старых, их взаимоотношения, но то, что одни формы происходят от других, остается неизменной основой объяснений. Усилия исследователей направлены на то, чтобы понять, как, в каком процессе возникло "ошеломляющее разнообразие жизни". Интерпретаторы таксономии считают, что когда-то жившие "просто устроенные" таксоны дали начало современным "сложно устроенным" таксонам. Таксоны произошли от таксонов. Само отношение "происходить от" при его эволюционной интерпретации считается вполне определенным, простым в том смысле, что оно не нуждается в теоретическом анализе и дополнительном осмыслении. Однако это далеко не так. Более того, экспликация содержания отношения "происходить от" позволяет считать его применение в эволюционных учениях неверным в принципе. Отношение "происходить от" эмпирически определено в таких областях знания, как антропология (и структурная антропология в особенности), этнография, а также в практике семенного и племенного дела. Это отношение означает, что некоторым особям можно сопоставить других особей в качестве потомков или предков. Структуры родства имеют сетчатый характер, в которых каждый член рассматриваемой популяции (племени, клана) находится в родстве с каждым другим членом. Естественно, что степени родства варьируют и их каталогизация является, в частности, одной из задач генетики популяций. И.И. Шмальгаузен утверждает, что сетчатое родство существует только до тех пор, пока продолжаются скрещивания, т.е. пока новые формы не вышли за пределы вида90. Это означает, что за пределами круга особей, скрещивающихся друг с другом, если и можно установить родство, то какое-то такое, которое не связано со скрещиванием. Что же это за родство? Из антропологии известно, что и в так называемых примитивных, и в цивилизованных культурах на биологическую структуру родства, имеющую сетчатый характер, накладываются собственно социально-экономические интерпретации, при которых одни биологические отношения считаются существенными (в праве наследования, например), в то время как другие при установлении родства не принимаются во внимание. Данные этнографии и структурной антропологии дают основание различать и противопоставлять биологические основания родства и их социальные интерпретации, носящие, как правило, мифологизированный характер91. При социально-экономической интерпретации отношений родства и происхождения формируется генеалогическое древо, которое определяет, от каких предков произошли представители династии, рода, семьи и, следовательно, на какой социальный статус могут рассчитывать потомки этих предков. Эмпирическое содержание генеалогического древа минимально, так как линейное биологическое родство из-за сетчатости его структур и зависимости от достаточно ситуативных институализаций межполовых отношений установить невозможно. Однако мифологизированный компонент генеалогического дерева устанавливает линейное происхождение и выделяет ключевых предков, что и определяет социальный статус потомков. Вполне однозначно отношение "происходить от" определено только в практической деятельности людей - в получении чистых линий лабораторных животных, выращивании культур микроорганизмов и т.д. Проблема происхождения в этих областях практики решается созданием условий для скрещивания или других форм размножения и строгой фиксацией этих условий, что дает возможность точно знать, от какой особи или группы особей ведет род конкретное животное, растение, культура микроорганизмов. Строго говоря, деятельность по получению чистых линий представляет собой аналитическую работу, основанную на эксперименте и соответствующих типологических представлениях. Совокупность абстракций, т.е. начальных условий для получения чистых линий, еще нуждается в особом анализе. Отношение "происходить от", таким образом более или менее строго определено только как отношение между особями и популяциями особей. Родство и происхождение как популяционные отношения изучаются в настоящее время совокупностью экологических областей знания, где, кроме прочих, рассматриваются и проблемы взаимосвязи популяций, то, как они переходят друг в друга, изменяясь при этом до неузнаваемости92. Однако при эволюционной интерпретации популяционных отношений возникает своеобразная и парадоксальная ситуация: отношения происхождения и родства исследуются как отношения между особями и популяциями, а интерпретируются как отношения между таксонами в иерархии (или другой системе) таксонов. Таксоны, как это отмечалось ранее, формируются исследователями в рамках таксономического цикла исследования при установлении между особями отношений сходства. Значение таксонов и их систем, упорядоченных биологическими определителями, прежде всего в том, что они позволяют ориентироваться в существующем и существовавшем многообразии форм жизни, свертывают многообразие и структуру, знание об отношениях в которой и умение пользоваться определителями дают возможность любому специалисту отнести найденную и описанную по определенным правилам особь к какому-либо таксону. Однако полезность классификационных систем не считается достаточным основанием для оправдания их существования93. Структура, предназначенная для упорядочения форм жизни, по мнению систематиков, должна еще и отображать процесс, в котором многообразие сформировалось. Усилия интерпретаторов классификационных систем как филогений направлены на то, чтобы определить, как и каким образом, а может быть, и когда таксон Х произошел от таксона Y. При этом отношения сходства, в которых только и существуют таксоны, интерпретируются как отношения родства94. Наиболее распространенные варианты эволюционного учения - дарвинизм, номогенез, ламаркизм - сходны в том, что они устанавливают между таксонами отношения родства и происхождения, но коренным образом расходятся в том, как эти отношения проявляются в процессе эволюции. Дарвинисты считают, что многообразие форм жизни произошло в процессе дивергенции сходных признаков, в то время как номогенетики стоят на том, что эволюцию определяет процесс конвергенции признаков. Ламаркисты постулируют приспособительные прижизненные изменения, появление новых признаков у таксонов и их наследование. При всем различии в интерпретации путей и направлений эволюционного процесса существующие и возможные варианты эволюционных учений, как показал А.А. Любищев95, обладают некоторой инвариантной структурой, позволяющей комбинировать исходные посылки таким образом, что практически любую эволюционную схему можно представить в виде исходных оппозиций и отношений между ними. Основатель структурной антропологии К. Леви-Стросс96, много внимания уделивший изучению мифологических интерпретаций отношений родства и происхождения, в своих работах также использовал аппарат, позволивший ему свести многообразие известных мифов в некую инвариантную структуру. Конечно, сходство еще далеко не родство, но в данном случае оно позволяет объединить мифы, изученные К. Леви-Строссом, и варианты эволюционного учения в один таксон - мифологемы, имеющие сугубо социальное происхождение. Представляется, что отношение между реальными процессами, в которых сформировалось и формируется многообразие форм жизни, и их интерпретациями в эволюционных учениях примерно такое же, как между биологическими основаниями родства и генеалогическими древами. И в том и в другом случае на совокупность особей, связанных отношениями родства и происхождения, накладывается интерпретационная схема, отражающая доминирующий в культуре способ осмысления реальности и связанная с субстратом, в котором отношения родства и происхождения реализуются, очень опосредствованно, причем опосредование не научно, не требует эмпирической проверки, а принимается на веру. В мифологических интерпретациях иерархии таксонов обращает на себя внимание разительное противоречие между конкретными наблюдениями и экспериментами и их эволюционными толкованиями. В биологии, да и в любой другой описывающей науке, превращение таксона в таксон, т.е. нахождение в потомстве одной особи экземпляров, могущих быть отнесенными к другому таксону, никогда не наблюдалось. Более того, культурные растения, совершенно не похожие на своих предков, относятся тем не менее к тому же таксону, что и предки, они считаются не более чем разновидностями исходного вида. П. и Дж. Медавары в работе, имеющей подзаголовок "Современные концепции в биологии", пишут: "Никаких причудливых гетерогенезов никогда не случается, вопреки всевозможным экстравагантным представлениям, когда эмпирическая точность не считалась необходимым или хотя бы желательным качеством претендующего на точность изложения"97. И несколькими абзацами ниже эти же авторы пишут: "...время от времени возникают отклонения, которые задним числом можно рассматривать как источник новых видовых форм"98. Когда деятели от биологии под руководством Т.Д. Лысенко доказывали, что в потомстве кукушки может появиться пеночка, из овса может вырасти овсюг, а вирусы самопроизвольно превращаются в бактерии99, это вызвало негативную реакцию ученых, и в дальнейшем исповедование таких взглядов стало несовместимым со статусом биолога. Однако невозможное сейчас, никем никогда не наблюдавшееся теми же самыми учеными, которые выступали против догм неоламаркизма, считается возможным в геологические времена. Например, И.И. Шмальгаузен считал, что в процессе эволюции, при постоянном расхождении признаков, увеличивается многообразие органических форм и что одного общего корня возникли в геологическом прошлом крупнейшие подразделения растительного и животного царства, представители которых постепенно усложнялись, повышали свою организацию и дифференцировались все дальше, давая начало новым подразделениям100. Легенда о золотом веке, в котором ныне невозможное и невероятное осуществлялось с необходимостью, осталась, похоже, только в эволюционных мифологемах, в которых геологическое время имеет существенное сходство с мифологическим временем. В мифологическом времени эволюционных учений таксонам приписывается тот же модус существования, что и особям. Таксоны в нем рождаются, живут и умирают-вымирают. Признаки таксонов могут сходиться и расходиться, формируя новые таксоны, как пары в бальных танцах. Наверное, не случайно содержание дискуссий по проблемам эволюционной интерпретации иерархии таксонов сходно с содержанием средневековых споров о реальности универсалий, что отмечается и некоторыми методологами биологии. Такова сила представлений, сформированных много веков назад, в эпоху, когда проблемы родства, происхождения, генеалогии были социально актуальны. Эти представления оформились как мифологема биологии во внутренне связную систему, независимую от эмпирии, более 150 лет назад в учениях Ч. Дарвина, К. Ламарка и других эволюционистов и продолжают свое существование сейчас. Уже несколько десятилетий существуют области науки, эмпирически и теоретически исследующие процессы, в которых одни популяции превращаются в другие, как при этом меняются физиологические, генетические, этологические и другие свойства особей101. Но результаты этих областей науки, в принципе самодостаточные, в мифологических интерпретациях иерархии таксонов используются для доказательства или опровержения того или иного варианта мифологемы. Разрыв между мифологемами и собственно исследованиями настолько велик, что соотнести результаты науки с эволюционными интерпретациями практически невозможно. Исследования идут сами по себе, описываются все новые и новые формы жизни, усовершенствуются ключи-определители, становятся более или менее ясными реальные пути и направления эволюционного процесса, а мифологемы существуют в неизменном виде. Уже описанное и упорядоченное ключами многообразие форм жизни настолько богато, что позволяет получить выборки наблюдений и фактов, служащих для обоснования самых экстравагантных вариаций учения о происхождении одних таксонов от других. Итак, задачей таксономии являются описание многообразия особей (отдельностей) и упорядочение этого многообразия. Последнее достигается в построении системы таксонов, основанной на констатации сходств между описанными представителями многообразия. Система таксонов имеет, как правило, форму иерархии, хотя для бактерий, например, иерархические классификации оказываются непригодными102. Сходство между особями неправомерно интерпретируется как родство между таксонами. Последнее служит для наполнения системы таксонов филогенетическим содержанием, что в принципе неправомерно. Отказ от филогенетической интерпретации иерархии чреват для биологии, как пишет Л.П. Татаринов103, потерей идеологического содержания, интепретационной схемы, в которой биологам представляется осмысленной известная из практики упорядоченность форм жизни. Необходимость отказаться от представлений о том, что структура, в которой упорядочены формы жизни, отражает ход и результаты эволюции, вызывает у биологов негативизм прежде всего потому, что эволюционным мифологемам нет замены в их роли объясняющих построений таксономии. Разделение и некоторое противопоставление совмещенных в мифологемах аспектов происхождения и упорядочения форм жизни в современной биологии осуществляются несколькими путями, иногда паллиативными, иногда достаточно революционными. В частности, есть работы, в которых отношения родства и происхождения исследуются эмпирически и теоретически, но интерпретируются как отношения между таксонами ранга вида, причем таких видов, которые можно однозначно сопоставить с популяциями104. Виды тем самым получают выделенный статус среди других таксонов, и многие методологи биологии считают, что эволюционируют только виды, в то время как таксоны более высоких рангов, если и эволюционируют, то каким-то другим образом, нежели виды105. Но противопоставление микро- и макроэволюций оправдано только тем, что эволюционные процессы протекают на уровне особей и популяций, иногда принадлежащих одному виду (т.е. относимых исследователями к одному виду), но интерпретируются как эволюция видов. Распространение знаний об эволюции популяций на таксоны более высоких рангов, чем вид, вызывает логические трудности даже у самых правоверных эволюционистов. Новое вино - знание об эволюции популяций - пытаются влить в старые мехи, поддержать видимость жизни в средневековом мировоззрении. Гораздо более обоснованным представляется подход к иерархии, полностью разводящий сходство форм жизни, служащее для конструирования таксономических систем, происхождение и эволюцию. Со времен И.В. Гете существует представление, что сходство особей может иметь совершенно иные, не филогенетические интерпретации. Снова возникает идея архетипа, которая сходство форм жизни связывает не с родством и происхождением, а с планом строения, т.е. совокупностью аналитических объектов, упорядоченных какими-то гипотетическими отношениями. Представления об архетипе пока существуют скорее как благие намерения, нежели конструкция, и больше обоснованы в методологической части, чем практической. В представлениях об архетипе таксономия в ее теоретической части пересекается с аналитическими науками, а не с эволюционными представлениями и концепциями. И если действительно объяснение в таксономии связано с архетипами (идеальными объектами, теоретическими мирами), то иерархия таксонов (или другие системы) не нуждаются ни в каких надстройках. Они сами по себе представляют теоретические структуры, выполняющие вполне определенные описывающие и упорядочивающие функции, что более чем достаточно, чтобы не искать им иных обоснований существования. Примечания:
|
|
||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||